
 0
0


1. Introducere
Artificializarea interrelaţiilor din cadrul biocenozelor agricole şi silvice, influenţa negativă a unor factori climatici, produc dereglări în lanţurile trofice, care favorizează înmulţiri populaţionale ale insectelor dăunătoare. Reducerea densităţii dăunătorilor nu se poate realiza fără utilizarea unei game largi de insecticide, preparate biologice (biopesticide) etc.
În prezent, în România sunt înregistrate peste 350 de molecule (substanţe active), formulate în peste 800 de produse comerciale, inclusiv amestecuri, utilizate atât în agricultură, cât şi în silvicultură (Berca, 2003). De asemenea, dezvoltarea durabilă din ambele domenii prevede, pe lângă eliminarea utilizării insecticidelor din grupele cu toxicitate ridicată, cu impact negativ asupra faunei utile, animalelor şi omului şi reducerea numărului de substanţe active folosite. Prin utilizarea durabilă a pesticidelor, concept elaborat recent în cadrul Uniunii Europene, se prevăd analize minuţioase ale preparatelor în vederea omologării, reducerea cantităţilor folosite la unitatea de suprafaţă, precum şi a numărului de tratamente. Urmare a acestor evoluţii conceptuale, în managementul integrat al dăunătorilor o mare importanţă are elaborarea unor tehnologii în care produsele de combatere să fie utilizate după modul de acţiune, pentru siguranţa folosirii şi pentru evitarea apariţiei rezistenţelor încrucişate. În acest scop, lucrarea de faţă face o sinteză a modului de acţiune al preparatelor, pentru o înţelegere mai bună a fenomenelor care pot să apară în practica de zi cu zi a specialiştilor din domeniul protecţiei plantelor.
2. Modul general de acţiuneUn produs de combatere acţionează prin diferite interacţiuni moleculare cu un anumit tip de proteină. Corpul insectelor, conform eredităţii, formează multe tipuri de proteine, care alcătuiesc ţesuturi, organe, exoscheletul insectei etc. Alte proteine joacă rolul unor catalizatori, ai unor reacţii chimice, în depozitarea sau transportarea produselor nutritive sau a celor provenite din dezasimilaţie, în mişcarea unor molecule prin membranele celulare.
Majoritatea insecticidelor, acaricidelor sau unii metaboliţi (biopesticide) acţionează pe proteine-ţintă cuprinse în sistemul nervos semnalizator (prin perturbarea receptorilor neurotransmiţători), participante în respiraţia celulară (în calitate de perturbatori ai respiraţiei) sau pe cele care au rol în creştere şi dezvoltarea insectelor (în perturbarea activităţii regulatorilor de creştere) (Salgado, 1997). Insecticidele se pot lega de proteine pe unul sau mai multe locuri ţintă, cu acţiuni de activare sau inhibare ale funcţiunilor, cu efecte perturbatorii, ale căror simptome se numesc mecanisme de acţiune.
În funcţie de modul de legare pe proteina-ţintă, se stabileşte modul de acţiune al produsului de combatere, se determină selecti-vitatea în folosire, viteza de acţiune şi rezistenţa gazdei.
Întrucât organismele animale conţin multe proteine asemănătoare, cu funcţie identică, există riscul ca produsele să acţioneze asupra proteinei-ţintă, însă atât asupra insectelor dăunătoare, cât şi a faunei utile şi a omului.
3. Modul de acţiune a insecticidelor 3.1 Acţiunea asupra sistemului nervos.Sistemul nervos al insectelor este format dintr-o reţea complexă de neuroni, care transmite informaţiile de la senzorii externi (ce recepţionează diferiţi factori biotici şi abiotici) şi de la cei interni (receptori pentru funcţionarea diferitelor organe şi sisteme, parametri biochimici ai proceselor fiziologice etc.) la sistemul nervos central. Toate aceste semnale se propagă sub formă de impulsuri nervoase (potenţial de acţiune), de natură electrică, prin axonul neuronilor (care formează nervii), legaţi cu dendritele neuronilor vecini prin joncţiuni specializate, numite sinapse. Aglomerările de neuroni ce formează ganglionii, respectiv sistemul nervos al insectei, sunt legate de organele senzoriale externe sau interne, prin nervi senzitivi, care transmit informaţiile, şi prin structuri efectore, formate din nervii motorii.
Modul de acţiune al receptorilor neurotransmiţători, care controlează fluxurile de ioni prin canale transmembranice, aşa numiţi receptori ionotropici, precum şi perturbările provocate de insecticide sunt prezentate în tabelul 1 (Salgado, 1997; Bloomquist, 1996; Haulică et al., 2002).
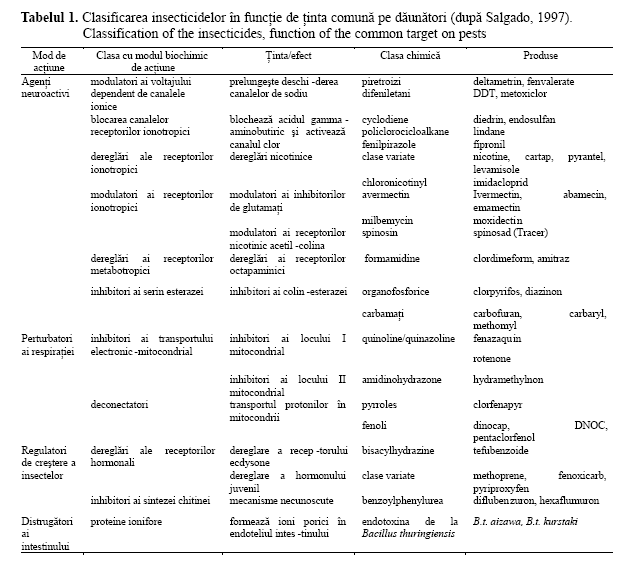
A. Acţiunea asupra axonului. Deoarece neuronii sunt separaţi de lichidul extracelular printr-o membrană celulară, impulsul nervos (potenţialul de acţiune) este transmis transmembranic prin fluxuri de sarcini. Diferenţa de potenţial este generată de acţiunea a doi curenţi ionici puternici. Ionii de potasiu (K+) sunt de zece ori mai abundenţi în interiorul celulei decât în afara ei, iar în repaus, membrana este slab permeabilă la aceştia. Pierderea ionilor de potasiu încărcaţi pozitiv (K+), prin părăsirea celulei, o încarcă negativ şi, totodată, opresc scurgerea acestora. În faza de repaus, se poate spune că celulele au un potenţial negativ (-50 la-100 mV). Ionii de sodiu (Na+) sunt de zece ori mai abundenţi în afară celulei decât în interiorul ei, iar negativitatea acesteia creează o forţă electrochimică puternică, atrăgându-i în celulă. Acest lucru nu se întâmplă în faza de repaus, deoarece membrana celulară le este puţin permeabilă.
Membrana conţine o concentrare puternică de proteine, numite canale electrice dependente de sodiu. Inactive în stare de repaus, acestea simt potenţialul de acţiune transmembranic şi se deschid, atunci când devin pozitive. Dacă un semnal (impuls nervos) intră printr-o terminaţie a axonului, formează în membrană un potenţial pozitiv, care deschide numeroase canale pentru ionii de sodiu. Astfel, dacă starea de repaus potenţial înseamnă o concentraţie puternică de ioni K+ în interiorul celulei şi o slabă permeabilitate a membranei la aceştia, acţiunea potenţială este dată de o concentrare externă puternică de ioni de Na+ şi de o puternică permeabilitate a membranei pentru aceşti ioni. Cele două procese intră în joc, determină acţiunea potenţială şi restabilesc pauza de negativitate din celulă. Restabilirea stării de negativitate din interiorul celulelor se face prin oprirea influxului ionilor de Na+ şi prin deschiderea canalelor electrice dependente de K+, ce permit ieşirea ionilor şi restabilirea sarcinii negative din celule.
Insecticidele selective pentru sistemul nervos al insectei interferează cu transmisia semnalelor electrice prin axon. DDT-ul şi piretroizii se leagă de proteinele canalelor de sodiu, care le pot închide sau activa. Acestea, chiar în cantităţi foarte mici pe canalele de sodiu, le menţin deschise pentru o nouă acţiune potenţială, cu un influx de ioni de Na+, generând şi multiplicând aceste acţiuni potenţiale. Canalele rămân deschise şi impulsurile reexcită de mii de ori aceeaşi secţiune de membrană. Prin aceste dereglări funcţionale, sistemul nervos al insectei este excitat continuu de impulsurile nervoase, fapt care conduce la oboseala musculară prin contractării repetate ale muşchilor, urmate de paralizia şi moartea insectei.
B. Acţiunea asupra sinapsei. În cadrul sinapsei, două celule sunt separate de un spaţiu de 200-300A, cunoscut şi sub numele de fisură sinaptică. Semnalul este transmis de la o celulă la alta, pe cale chimică, prin aşa numiţii neurotransmiţători.
B.1 Acetilcolina ester al acidului acetic şi colina. La o acţiune potenţială, în terminaţia presinaptică, se declanşează fuziunea veziculelor sinaptice (mici pachete ce conţin acetilcolină) cu membrana celulară. Din fuziune rezultă expulzarea Ach în spaţiul sinaptic. Moleculele de Ach difuzează prin membrana postsinaptică, unde se leagă de receptorii nicotinic Ach. Când două molecule de Ach se leagă de o moleculă nicotinică, ele deschid canalele, prin care ionii de sodiu pot pătrunde în celula postsinaptică, ducând la depolarizare şi excitare.
Asemănător axonilor, acţiunea potenţială se termină prin închiderea canalelor de sodiu. Procesul are loc rapid, deoarece o concentraţie mare de acetilcolinesterază o desface în compuşii acid acetic şi colină, care sunt transportaţi înapoi în celulele presinaptice şi reciclate.
Insecticidele intervin în acţiunea receptorului nicotinic Ach şi acetilcolinesteraza. Imidaclopridul şi alte insecticide nicotinice imită acţiunea Ach pe receptor, producându-le perturbări funcţionale.
La insecticidele descoperite în ultimii 30 de ani (organofosforice şi carbamaţi), locul ţintă este acetilcolinesteraza, de care se leagă catalitic şi, în acest mod, previne desfacerea acetilcolinei, prelungindu-i acţiunea în celula postsinaptică.
B.2 Acţiunea asupra altor neurotransmiţători. Pe lângă acetilcolină, în organismul insectelor se găsesc şi alţi neurotransmiţători, fiecare cu specificul sintezei, recepţiei şi inactivării lor. În combaterea insectelor are importanţă şi acţiunea insecticidelor privind neurotransmiţătorii: acidul gamma-aminobutiric (GABA), glutamat şi octopamină. În contrast cu receptorii Ach, receptorii GABA şi inhibitorii de glutamat activează canalele ce permit curgerea în celulă a ionilor de clor încărcaţi negativ. Influxul de încărcătură negativă inhibă activitatea neuronilor şi neutralizează activitatea Ach. Unele insecticide (dieldrin, endosulfan, lindan, fipronil) blochează activitatea GABA şi excitaţia nervoasă se răspândeşte în tot corpul. Inhibarea receptorilor glutamate este strâns legată de activitatea GABA şi conduce la inhibiţie totală şi paralizie (avermectin, milbemycin).
În contrast cu receptorii ionotropici, receptorii metabotropici transmit mesajul intracelular prin stimularea enzimelor. De exemplu, în organul luminiscent al organismelor, sub acţiunea octapaminei (produsă de neuroni) se formează adenozin monofosfat, care activează enzimele producătoare de emitere chimică a luminii. Anumite insecticide şi acaricide formamidine (clordimeform, amitraz) imită acţiunea receptorilor octopaminici, care se găsesc în sistemul nervos central al insectei, creează hiperexcitabilitate şi chiar moartea insectei.
3.2 Perturbarea procesului respiratorCelulele au nevoie de energie, care se obţine din carbohidrazele, lipidele şi aminoacizii din hrană. Energia este stocată prin formarea unui intermediar bogat în energie, adenozintrifosfat (ATP), care aprovizionează procesele metabolice celulare. Transformarea energiei din hrană în ATP se petrece pe suprafaţa mitocondriilor, ce prezintă un lanţ de enzime, care captează energia rezultată din oxidarea controlată a carbohidrazelor, grăsimilor şi aminoacizilor. În oxidare se foloseşte oxigenul molecular, procesul numindu-se şi respiraţie mitocondrială. Energia chimică rezultată din oxidare pompează ionii de hidrogen, încărcaţi pozitiv, în afara mitocondriei, în celulele înconjurătoare, lăsând interiorul acesteia negativ. Energia rezultată din hrană este stocată temporar sub o formă de voltaj, care străbate membrana mitocondriei. Enzima ATPsintetaza (din membrana mitocondrială) permite întoarcerea ionilor de hidrogen în celulă, folosind energia eliberată din rezultatul neutralizării sarcinii, prin sinteza de ATP.
Insecticidele pot perturba respiraţia mitocondrială pe două căi. Prin inhibarea transportului electronilor mitocondriali ai lanţului, pe locul I (rotenona, fenzaquin) sau pe locul II (hydramethylon). Alte insecticide pot acţiona ca decuplatori între producţia de ATP şi transportul ionilor de hidrogen (insecticidele chlorfenapyr şi acaricidele fenolice).
3.3. Regulatorii de creştere a insectelor
Asemănător organismelor cu schelet extern, creşterea insectelor se realizează prin năpârlire, formându-se o nouă cuticulă. Anumite insecticide inhibă formarea unei noi cuticule, dar nu se cunosc locurile ţintă de care se leagă aceste produse. Formarea noului înveliş este controlat şi de hormonul năpârlirii.
În dezvoltare un rol important îl are hormonul juvenil, prezent la năpârliri, mai puţin la stadiul de adult. Juvenoidul sintetic methoprene, imită pe cel natural şi creează o structură anormală între larvă şi adult, dar care nu se poate reproduce şi moare imediat.
4. Modul de acţiune a preparatelor biologiceSunt cuprinse modurile de acţiune a bacteriilor, baculovirusurilor, ciupercilor şi metaboliţilor acestora.
4.1 Bacteriile. În general, bolile produse de bacterii se caracterizează prin pătrunderea în hemocel şi multiplicarea patogenului, ca urmare insecta murind prin septicemie (Galani şi Andrei, 1997).
Tulpinile bacteriei Bacillus thuringiensis, care se foloseşte cel mai mult în combaterea biologică, produc endotoxine cu caracter insecticid. Acestea se leagă de proteinele din intestinul mediu al insectei. Ajunse în membrană formează canale, care permit scurgerea ionilor şi distrugerea celulelor. Producerea acestor toxine se realizează şi prin inginerie genetică, prin introducerea genelor producătoare de endotoxine în genomul unor plante, rezultând organisme modificate genetic.
4.2 Baculovirusurile. Sunt vibrioni asamblaţi în formaţiuni proteice numite supraviriocapsizi (SPVC) pătrund în corpul insectelor prin ingestie, ajung în intestinul mediu şi, sub acţiunea enzimelor, se proteinizează şi eliberează vibrionii, care se dezvoltă în celulele epiteliale şi pe care le distrug (Ciuhri, 1997).
4.3 Ciupercile. Pentru producerea unei micoze, patogenul fungic trebuie să penetreze gazda printr-o acţiune mecanică şi una de natură enzimatică. După străpungerea tegumentului, ciupercile pătrund în hemocel, unde se multiplică. Colonizarea insectei se face prin proliferarea blastoporilor şi creşterea continuă a miceliului, care blochează circulaţia sanguină şi dezintegrează total ţesuturile.
În ce priveşte modul de acţiune asupra sistemului nervos, se prezentă spinosadul, un metabolit natural obţinut din actinomiceta Saccharopolyspora spinosa, care deschide o nouă clasă de produse naturale derivate din organisme (Bret et al., 1997, Peterson et al., 1997, Salgado, 1997, Thompson et al., 1997). Spinosadul se leagă de una sau mai multe proteine ale sistemului nervos. Faţă de alte produse, acţionează pe un loc diferit pe receptorii nicotinici, iar, prin activarea receptorilor nicotinic-acetilcolină, produce un influx de ioni de sodiu, care depolarizează neuronii ce devin hiperactivi, excitând musculatura corpului (manifestată prin extensia picioarelor, tremur, bătăi din aripi, înghiţire de aer). Se produce o oboseală neuromusculară din care rezultă paralizia insectelor. Prin descoperirea unor astfel de biopesticide, cu un mod diferit de acţiune, se pot înlocui unele insecticide mai toxice pentru ecosisteme şi se evită apariţia rezistenţei încrucişate.
5. Discuţii şi concluziiSe poate constata că preparatele, fie chimice, fie biologice, acţionează prin legarea pe anumite proteine-ţintă, care perturbează transmisia semnalelor prin sistemul nervos, respiraţia mitocondrială, digestia în intestinul mediu, precum şi creşterea şi dezvoltarea insectelor.
Pentru prevenirea producerii fenomenelor de rezistenţă, de mare importanţă este utilizarea raţională a produselor care acţionează pe aceeaşi proteină-ţintă. De exemplu, se apreciază că majoritatea insecticidelor organofosforice şi carbamice acţionează asupra ţintei acetilcolinesteraza. Deci, cu toată diversitatea acestora, folosirea preparatelor cu aceeaşi proteină-ţintă, va mări riscul apariţiei de rezistenţă încrucişată. Pentru evitarea apariţiei fenomenului de rezistenţă sau a rezistenţei încrucişate trebuie evitată folosirea produselor din aceeaşi clasă chimică pe perioade lungi de timp, deoarece ele acţionează pe aceeaşi proteinăţintă. Rotaţia preparatelor din diferite grupe chimice, cu diferite moduri de acţiune, asigură o combatere eficientă.
Prin descoperirea a noi clase de produse naturale, cu acţiune insecticidă, cu mecanisme diferite privind legarea de proteinaţintă şi a modurilor lor de acţiune (vezi spinosadul), se elimină apariţia fenomenului de rezistenţă încrucişată şi se asigură folosirea acestora pe perioade lungi de timp.
Referitor la rezistenţa insectelor la diferite insecticide, acesta se dezvoltă prin patru mecanisme: (i) modificarea comportamentului insectelor prin evitarea consumului hranei tratate sau a contactului cu insecticide prin fenomenul de repelenţă; (ii) modificarea metabolismului, prin creşterea cantitativă şi cea a eficacităţii unor enzime care descompun insecticidele. Acest mecanism de rezistenţă metabolică poate da naştere la rezistenţă încrucişată; (iii) prin scăderea ratei de penetrare a insecticidelor, timp în care enzimele metabolice le descompun; (iv) prin mecanism de rezistenţă major, în care proteina-ţintă îşi modifică forma şi reduce acţiunea insecticidului. Acest tip de reacţie produce rezistenţă prin încrucişare la toate produsele din aceeaşi clasă chimică, de asemenea, şi la produsele care acţionează pe aceleaşi locuri ţintă. Un exemplu, rezistenţa la DDT, piretroide şi la dieldrin produce rezistenţă încrucişată şi la lindan, fipronil şi la toate ciclodienele (Salgado, 1997).
Din modurile de acţiune a insecticidelor sau preparatelor biologice se pot desprinde următoarele concluzii: (i) managementul combaterii integrate a dăunătorilor trebuie realizat de specialiştii din domeniul protecţiei plantelor, cu o pregătire corespunzătoare; (ii) în schemele de combatere se va evita folosirea repetată a produselor din aceeaşi clasă chimică sau chiar a altor clase chimice, dar care acţionează pe aceeaşi proteină-ţintă, pentru a evita rezistenţa încrucişată; (iii) respectarea dozelor de utilizare prescrise îndepărtează presiunea apariţiei fenomenului de rezistenţă la produse; (iv) cercetarea pune la dispoziţie noi produse naturale, cu un mod de acţiune diferit (spinosad), care elimină rezistenţa încrucişată şi au o toxicitate redusă faţă de mediu; (v) pentru realizarea dezideratelor de gestionare a unor ecosisteme durabile, eforturile cercetării trebuie îndreptate spre folosirea şi descoperirea de noi metaboliţi, cu caracter insecticid, şi cu un mod de acţiune deosebit, prin legarea de proteina-ţintă.
Bibliografie
Berca M., 2003. Mesajul preşedintelui Societăţii de Protecţia plantelor, Proplant 2003, Călimăneşti.
Bloomquist J.R., 1996. Ion channels as target for insecticides, Ann. Rev. Entomol., 41:163-190.
Bret B.L. et al., 1997- Biological properties of Spinosad, Down to earth, vol. 52, (1): 6-13.
Ciuhri M., 1997. Patologia insectelor şi lupta biologică, cap. din Ciochia V. şi colab., Limitarea populaţiilor de dăunători vegetali şi animali din culturile agricole prin mijloace biologice şi biotehnice în vederea protejării mediului înconjurător, Ed. Disz , Braşov, 491p.
Hăulică I. et al., 2002. Fiziologie umană, Editura Medicală, Bucureşti, 1369 p.
Peterson L.G. et al., 1997. Tracer naturalyte insect control and IPM, Down to earth, 52(1) :28- 34.
Price, N. R., 1991. Insect resistance to insecticides: mechanisms and diagnosis, Comp. Biochem. Physiol., 100C, pp. 319-326.
Salgado V. L., 1997. The modes of action of Spinosad and other insect control products, Down to earth, 52(1) :35-42.
Thompson G.D. et al., 1997. The discovery of Saccharopolyspora spinosa and a new class of insect control products. Down to earth, 52(1) :1-5.
Autorul: Conf. dr. Valentin Brudea-Facultatea de Silvicultură, Universitatea “Ştefan cel Mare”, Suceava. Poate fi contactat la adresa de e-mail: vbrudea@yahoo.com